 Meredith Root-Bernstein1
Meredith Root-Bernstein1Abstract
We carried out an experiment to understand the effect of reintroduced guanacos (Lama guanicoe, the wild relative of llamas) on the growth of the espino tree (Acacia caven), a common tree in the central Chilean espinal silvopastoral system. The guanacos browsed on the trees and stimulated growth. The guanacos also had several unexpected ecosystem functions in the espinal habitat. Reintroducing them to restore central Chilean woodlands seems very promising.
Introduction
This phase of the project formally ended in July 2016, and we are now starting on phase 2. We still have not had time to analyse all the data, but I can update you on our results and unexpected findings so far!
We started this project in order to test the hypothesis that guanaco browsing on the espino acacia would stimulate the trees to grow more than usual-- known as "compensatory growth". This hypothesis was based on similar observations in African acacias under browsing, and observations of the effects of guanaco browsing in lenga forests in the far south of Chile. However, since guanacos have not been in central Chile (except a few areas high up in the Andes) for around 500 years, and espinos have thorns, it was not clear whether or how this plant-herbivore interaction would work. We hoped that by stimulating espinos to grow, guanacos could be used as a natural restoration tool for the espinal habitat.
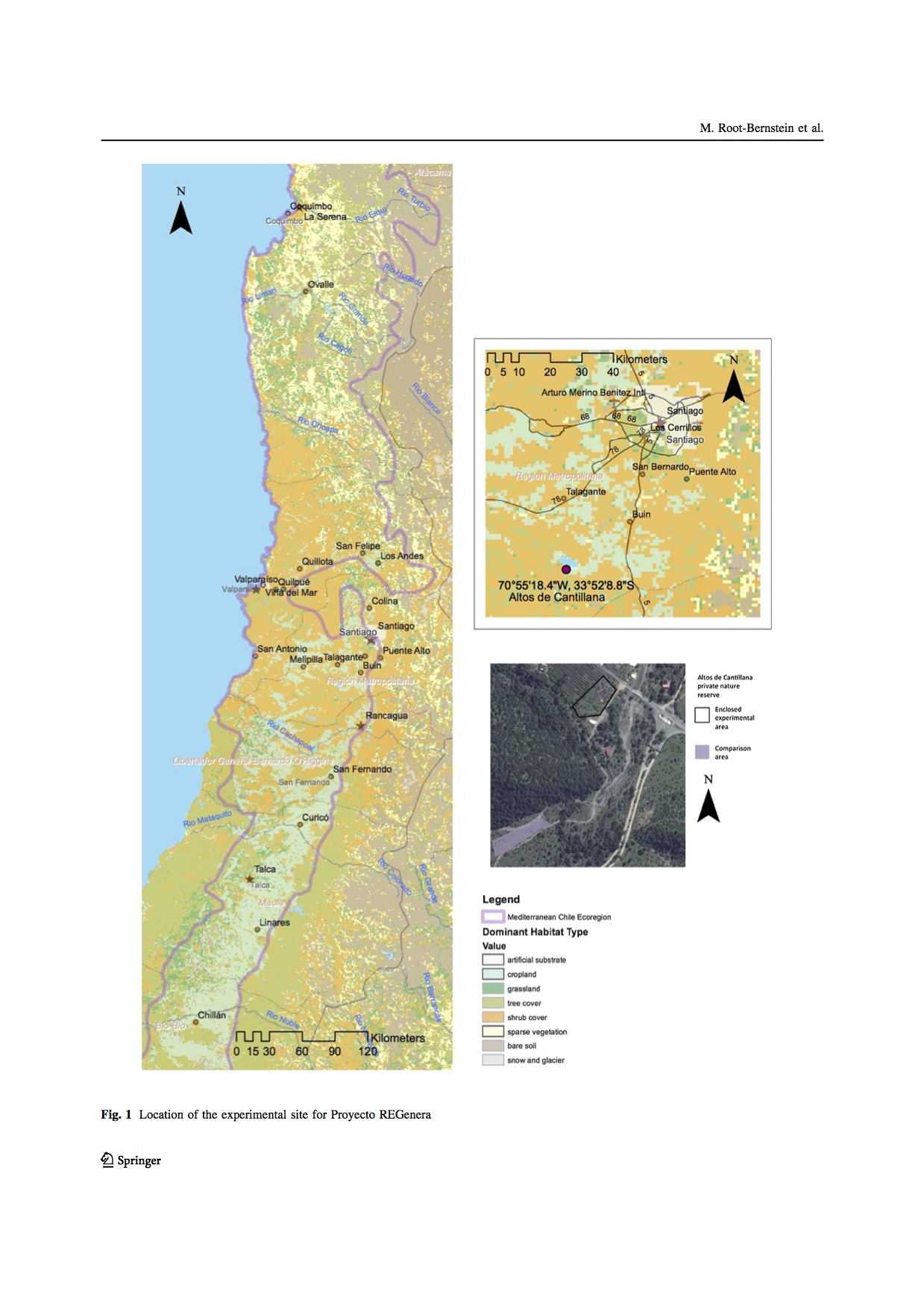
We were offered a loan of five castrated male guanacos from the Fauna Australis wildlife conservation unit at the Pontificia Universidad Católica de Chile. We were also offered a half-hectare espinal site in the Altos de Cantillana nature sanctuary. The owner had always dreamed of having guanacos on his land, but unfortunately this half-hectare of espinal was the only part that got any. We needed to build a fence with internal divisions in order to run the experiment and keep the guanacos safely, which was why we looked for funding through Experiment.
Methods
The fence had to be at least 1.8 meters high and surrounded by an electric fence, in order to keep out feral dogs and keep in the guanacos. The fence was made of wood and mesh, with wooden slats along the bottom to keep dogs from getting under it. We had some problems getting the project started because SAG, the agency in charge of animal welfare in this case, couldn't find time to inspect the site. But finally in June 2014 it was all ready:

I am quoting further methods from our paper, Root-Bernstein, M., Guerrero-Gatica, M., Piña, L., Bonacic, C., Svenning, J. C., & Jaksic, F. M. 2016. Rewilding-inspired transhumance for the restoration of semiarid silvopastoral systems in Chile. Regional Environmental Change, 1-16. In this paper, we present some preliminary results focusing on the guanaco's behaviour, as well as a spatial analysis (using GIS) of the potential connectivity between espinals to be gained from scaling up the guanaco reintroduction.
"The experimental site is a fenced 0.5 ha area of espinal atthe entrance to the private reserve Altos de Cantillana. The size of the site was dictated by the availabilityand feasibility of fencing the site. A fence surrounded byan electrical circuit was necessary not only to keep theguanacos inside the site, but also to keep out feral dogs.The guanacos used in the experiment are five castratedadult males from the breeding facility of Fauna Australis, aresearch group of the Pontifical Catholic University ofChile. Approval for the conditions of the experiment wasobtained from, and the ongoing welfare of the guanacos ismonitored by, SAG (Agriculture and Livestock Service)and the veterinarians on our research team.
"The site is divided into four sections of 0.125 ha each. Inwinter and spring (the wet season), the guanacos spendbetween 4 and 2 weeks in each section (the shorter timeinterval prevents over-grazing in late spring as aridityincreases) and are rotated from one section to the nextclockwise (the ‘‘pulse’’ treatment). In summer and fall(December–May), they are allowed to move freelythroughout the enclosure, in order to reduce the intensityof grazing on the herbaceous understory. Although Obaet al. (2000) report that Indigofera spinosa shows thelargest compensation effect when pruned in the dry season, we decided to implement the browsing pulse in thewet season due to the reverse phenology of A. cavencompared to other deciduous plants in Chile (it has leavesduring the dry season), which may also reverse its growthresponse relative to the season. In other words, we followOba et al. (2000) in implementing more-intense browsingwhen the trees have no leaves, in the expectation that thiswill stimulate increased growth during their next growthseason. Cromsigt and Kuijper (2011) also report thatearly-growth-season pulses of herbivory are associatedwith trees adapted to herbivory. Whether or not therotational plan replicates the natural migratory patterns ofguanaco in central Chile is unknown as we are not awareof any historical data on their migration in that area.Guanacos may have avoided colder temperatures at higheraltitude or further south by moving into the central valleyof central Chile in winter (the rainy season), where theycould have eaten the fresh herbaceous substrate andleafless A. caven. However, they may also have eaten A.caven leaves and shoots in early summer, before movingsouth or to higher elevations. Our rotational plan is basedon the expectation that winter herbivory will give thelargest compensatory growth response. The rotationalphase of the experiment also allows us to track the temporal lag of growth following known periods of herbivory.These data will help us to assess how long guanacosshould remain on a site to induce growth but not damagethe trees.
"The rotational plan...isintended to provide basic data on the guanaco–A. caveninteraction, including information on the conditions underwhich guanaco browse A. caven, which parts of the tree arebrowsed, the size of the growth response of A. caven toguanaco browsing, and the effects of season and browsingintensity on that response, that could be applied todesigning a transhumant rewilding system.
...
"The guanacos were introduced to Section A on June 18,2014. During the first month, we monitored their behaviorin order to determine an appropriate supplementary feedingamount. We currently lack the necessary data to calculatethe appropriate stocking rate for guanacos in espinal; thesedata will be obtained over the course of the experiment.The time required for the espinal to recover from intensiveherbivory is also unknown, so, given the restricted spaceavailable for the experiment, attenuating the herbivorypressure with supplementary alfalfa is necessary. Theamount of supplementary alfalfa is adjusted according tomonitoring of the guanacos’ body conditions.
"To determine how guanacos forage and behave in A.caven savanna habitat, we record behaviors using a focalanimal method. Each animal was observed by a singleindividual for four sessions of 10 min at a time, in a ran-dom order, over a period of 4 h during the day, 3 days perweek. Here, we report on observations in September,October, and November of 2014. Behaviors were recordedin JWatcher.
"In addition, we downloaded photographs from twocamera traps that were originally set up to monitor the siterather than guanaco behavior per se, but which capturedmany images of the guanacos. These were in sections A(October 2014, January, and April 2015), and B (Novem-ber 2014) of the enclosure. "
In addition, we collected the data on tree growth by photographing hundreds of individually marked branches, in both the experimental site and in a comparison site without guanacos. We photographed each marked branch once a month for 2 years, in three of the four sections each month, corresponding to the section currently with guanacos, the previous and the next sections in the rotation. These are the initial conditions:
| Section |
Enclosed area (ha) |
No. marked trees |
Mean tree height |
Mean tree canopy area |
Total marked branches |
| Experimental total |
0.49 |
||||
| Sector A |
0.125 |
64 |
2.3 ± 0.09 m * |
5.9 ± 1.15 m2 * |
136 |
| Sector B |
0.125 |
41 |
2.3 ± 0.17 m * |
7.2 ± 0.84 m2 |
213 |
| Sector C |
0.125 |
28 |
2.6 ± 0.17 m |
9.14 ± 1.40 m2 |
211 |
| Sector D |
0.125 |
26 |
2.6 ± 0.19 m |
8.25 ± 3.12 m2 |
209 |
| Comparison |
70 |
3.1 ± 0.11 m * |
12.7 ±1.22 m2 * |
198 |
Results
In the Root-Bernstein et al. 2016 paper, we report that the guanacos browsed on the espinos on most days, and that they spent some time eating branches and leaves independently of how much time they spent eating herbs and supplementary hay. When eating the leaves, they carefully picked the leaves out from between the spines with their long tongues, taking advantage of their narrow mouths. When eating the branches, they chewed the spines very carefully before swallowing. The more recently grown ends of the branches, despite having spines, should be relatively soft, and are the part of woody growth that browsers normally eat.
The browsed branches quickly grew back, as predicted. We also noticed that some small espinos were significantly damaged by the guanacos. They also grew some exceptionally long spines! However, we haven't yet been able to analyse the data on how much regrowth occurred after browsing.
One of the most interesting findings has not been reported yet. We were excited to discover the various ways in which guanacos alter their habitat. This included making trails, making dung middens, and making dust baths. This small-scale heterogeneity should provide niches for many herbaceous plants, insects, and their predators. In addition to its probable roles transporting seeds and affecting tree growth, these impacts might make the guanaco an ecosystem engineer.
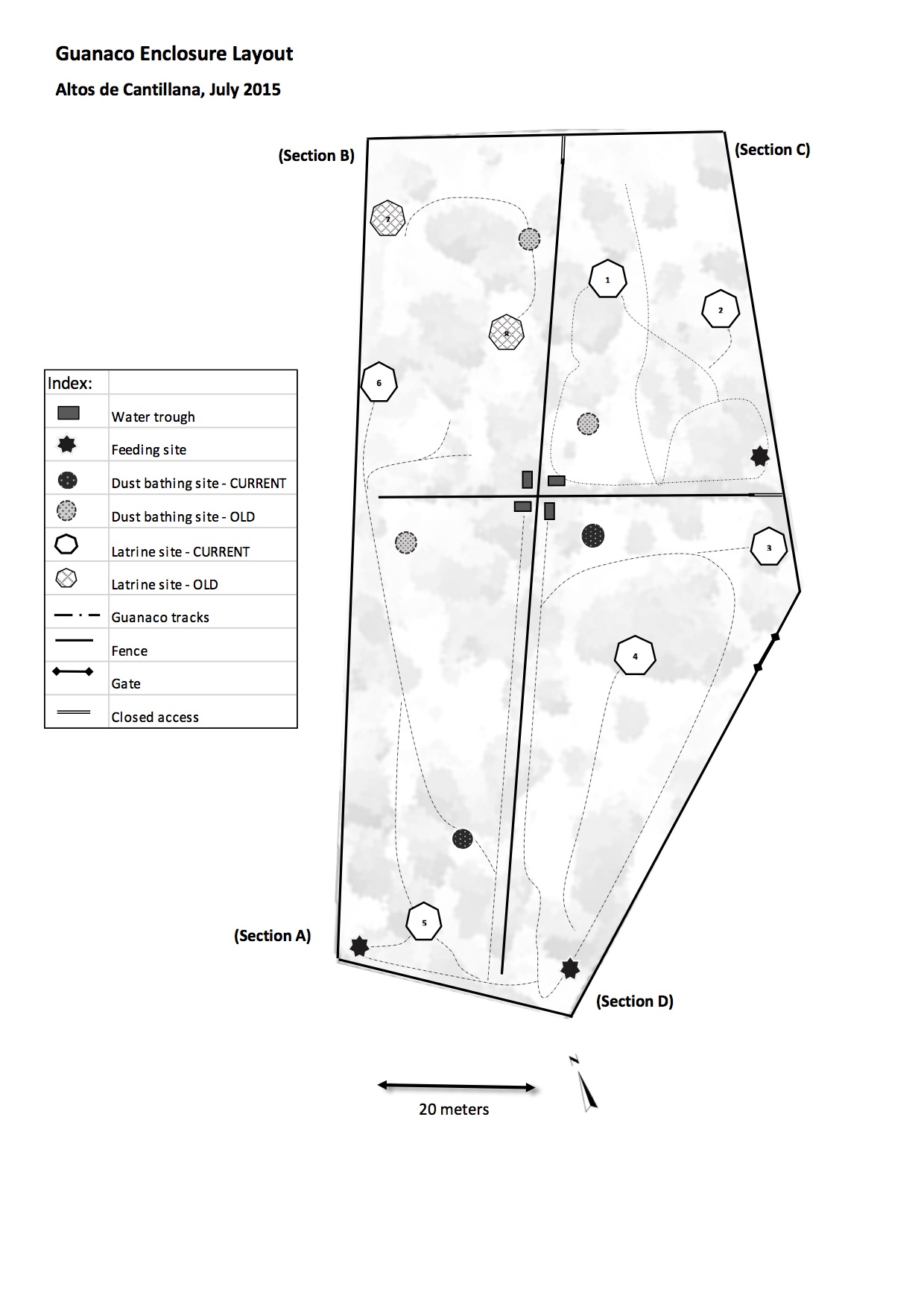
This is a map of the guanaco's constructions in 2015, made by volunteer Elise Dewar. Latrine = dung midden. The grey shapes are the espino trees.

We took photographs of the dung middens in each section of the enclosure each month. Here is an image from October 2015, where you can see how the dung middens provide extra nutrients for a range of herbaceous species, allowing them to grow unusually large. The California poppy that you can see with the orange flowers is not native to Chile, but is known to be able to thrive on high-nutrient soils. By taking up the nutrients and recycling them, the poppy is helping to turn the nutrients over to the soil in a form more available to native plants adapted to nutrient limitation-- so it is not necessarily bad that it is a nonnative species in this case. It would be interesting to known what native species used to have this niche or role when guanacos previously lived in this region.
Finally, we have also been doing some background research on this topic to support our future plans. In Root-Bernstein et al. (2016) and a forthcoming paper, we propose what we call "transhumant rewilding", or a kind of reintroduction-for-restoration in which the guanacos would not be totally wild. We propose this because extensive land cover change and habitat fragmentation in central Chile mean that a wild population of guanacos would run into all kinds of conflicts with humans and would probably be extirpated again (despite being a protected species in Chile). But, there is a long gradation of human-large herbivore relations between wild and domestic. We propose that guanacos could be herded between suitable habitat patches, similarly to how semi-wild reindeer have traditionally been herded in northern Europe and Siberia. We then used GIS analysis to ask what kinds of patterns of connectivity you could get between espinal patches that are in poor condition and need restoration, and espinal patches in good condition, under this model. Then, based on our research showing that espinal is successionally linked to sclerophyllous forest formation (Root-Bernstein et al. 2017), we extended this to see how the two woodland types could be linked by guanacos:
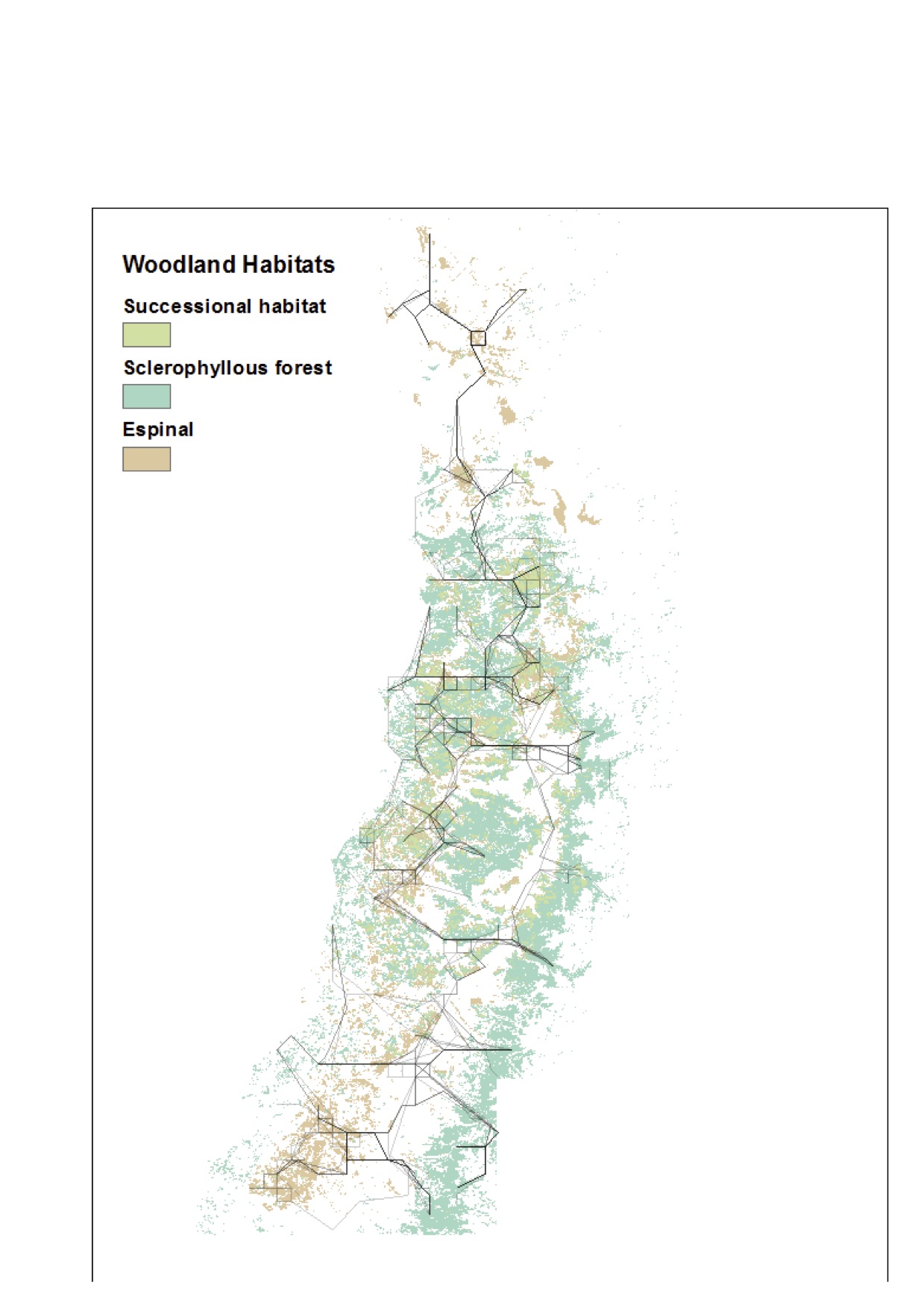
From our forthcoming paper, this figure shows the main pathways across central Chile, summed over years with different amounts of precipitation, along which guanacos could be herded to reach low-quality woodland from high-quality woodlands. This way, the guanacos could browse and transport seeds to areas that need restoration during winter, and graze and browse in richer habitats during summer. The figure shows that by targeting highest and lowest quality areas, the paths connect both types of woodland, as well as intermediate successional formations. This connectivity should increase woodland resilience and successional capacity (Newton & Cantarello 2015).
Conclusion
Next up, the guanacos will be transferred to the Cascada de las Animas nature sanctuary, where we have negotiated their release as wild animals. See our project to fund their release: https://experiment.com/project...
We hope to continue to monitor the successional processes and biodiversity of the dung middens, trails and dust baths, both at the original experimental site and where the guanacos will be released in Cascada de las Animas. We also hope to confirm that they are seed dispersers and how big of an impact they have on espino compensatory growth.
We have two specific predictions that were are working on finding additional funding or collaborations to test:
1) Guanacos may be seed dispersers for two rare endemic trees with relatively large seeds that currently have no known dispersal mechanism, the Belloto del norte, and the Chilean palm. Their rarity might be due to having lost their disperser-- can we conserve these rare and iconic trees with guanacos?
2) In Europe, European rabbits (which are invasive and very common in Chile) are facilitated by livestock which keep the grass short, the way rabbits like it. By contrast, smaller mammals around the world tend to be negatively impacted by large-hooved large herbivores like livestock, due to the risk of being trampled. When we replace the horses currently in Cascada de las Animas with guanacos, which graze less and stick to walking on trails more, we expect rabbits to decline and native small mammals to increase.











